
ARTÍCULO DE INVESTIGACIÓN
Efecto de la simbiosis micorrízica, la fertilización química y su combinación, en la relación suelo-planta del cultivo de morera
Effect of the mycorrhizal symbiosis, chemical fertilization and their combination, on the soil-plant relation of mulberry
Gertrudis Pentón1, R. Rivera2, G. J. Martín1, Aracelis Mena2, F. Alonso1 y Aida Medina2
1
Estación
Experimental de Pastos y Forrajes Indio Hatuey, Universidad
de Matanzas Camilo Cienfuegos, Ministerio de Educación Superior. Central
España Republicana, CP 44280, Matanzas, Cuba
2Instituto
Nacional de Ciencias Agrícolas, MES, Mayabeque, Cuba
Correo electrónico:
gertrudis@ihatuey.cu
RESUMEN
Se evaluó el efecto de la inoculación con hongos micorrízicos arbusculares (HMA) cepa Glomus cubense a través de canavalia (Canavalia ensiformis), la fertilización química y la combinación de ambas, en el funcionamiento radical, la simbiosis micorrízica y la capacidad de extracción de nitrógeno en la morera (Morus alba L.). Las dosis de fertilizante fueron: 1) testigo sin fertilización (F0); 2) 100-50-50 kg de N, P y K ha-1 por época (F1); y 3) 200-100-100 kg de N, P y K ha-1 por época (F2). Las alternativas para inocular HMA consistieron en: testigo sin HMA, aplicación directa al suelo, e inoculación a través de canavalia intercalada. La biomasa radical de la morera al inocular HMA a través de canavalia fue superior (490,28 mg 50 g-1 de suelo). La simbiosis micorrízica a los 150 días de aplicados los tratamientos fue superior con HMA inoculados a través de canavalia intercalada y complementada con F1 (375,33 esporas en 50 g de suelo y 1,46 % de densidad visual). La extracción de nitrógeno a través de la biomasa comestible, en el tratamiento HMA inoculados vía canavalia con F1, varió entre 245,68 y 336,75 kg ha-1 en la época de lluvia y entre 74,59 y 97,81 kg ha-1 en la poco lluviosa. Se concluye que la mayor distribución de raíces en el suelo, los mejores indicadores de simbiosis micorrízica y las mayores extracciones de nitrógeno de la morera estuvieron asociados de manera positiva a la inoculación con HMA a través de canavalia, complementada con fertilización química.
Palabras clave: biomasa, Canavalia ensiformis, hongos, inoculación.
ABSTRACT
The effect of the inoculation with arbuscular mycorhizal fungi (AMF) strain Glomus cubense through Canavalia ensiformis, chemical fertilization and their combination, on the root functioning, mycorrhizal symbiosis and the nitrogen extraction capacity in mulberry (Morus alba L.), was evaluated. The fertilizer doses were: 1) control without fertilization (F0); 2) 100-50-50 kg of N, P and K ha-1 per season (F1); and 3) 200-100-100 kg of N, P and K ha-1 per season (F2). The alternatives to inoculate AMF consisted in: control without AMF, direct application to the soil, and inoculation through intercropped C. ensiformis. The root biomass of mulberry when inoculating AMF through C. ensiformis was higher (490,28 mg 50 g-1 of soil). The mycorhizal symbiosis 150 days after the treatments were applied was higher with AMF inoculated through intercropped C. ensiformis and complemented with F1 (375,33 spores in 50 g of soil and 1,46 % of visual density). The nitrogen extraction through the edible biomass, in the treatment of AMF inoculated via C. ensiformis with F1, varied between 245,68 and 336,75 kg ha-1 in the rainy season and between 74,59 and 97,81 kg ha-1 in the dry season. It is concluded that the highest distribution of roots in the soil, the best indicators of mycorhizal symbiosis and the highest nitrogen extractions of mulberry were positively associated to the inoculation with AMF through C. ensiformis, complemented with chemical fertilization.
Key words: biomass, Canavalia ensiformis, fungi, inoculation
INTRODUCCIÓN
En la agricultura actual es necesario adoptar estrategias para estimular y optimizar procesos biológicos del suelo, que favorezcan el reciclaje de nutrientes y prioricen la adopción de técnicas multifuncionales que mantengan o mejoren la fertilidad, contrarresten los procesos de erosión y fomenten la presencia de poblaciones de organismos benéficos.
En tal contexto, los hongos micorrízicos arbusculares (HMA) se destacan dentro de la amplia gama de microorganismos del suelo que se utilizan en la producción de biofertilizantes (Calderón y González, 2007), cuya acción en la interfase suelo-raíz permite reducir las aplicaciones de fertilizantes minerales. Por ello, su empleo en los sistemas de explotación intensiva de la morera (Morus alba) para la producción de forraje (Martín et al., 2002; Villalba, 2009; Oropesa et al., 2011; Noda y Castañeda, 2012) podría ser una alternativa importante para reducir las altas cantidades de estos insumos en las condiciones actuales.
La inoculación de las plantas con hongos micorrizógenos induce el incremento de los procesos de absorción y translocación de nutrientes, tales como: P, N, K, Ca, Mg, Zn, Cu, Mo y B (Siqueira et al; 2010). Su efectividad se garantiza con prácticas de manejo que estimulan la multiplicación de propágulos infectivos (González et al., 2008).
El uso de la canavalia (Canavalia ensiformis) como medio de inoculación de los HMA constituye una vía para la biofertilización de plantaciones de morera establecidas, que a la vez puede ser complementada con diferentes fuentes y formas de fertilización (Martín, 2009).
Por todo lo anterior, el objetivo de esta investigación fue evaluar el efecto de la inoculación de los HMA a través de canavalia, la fertilización mineral y la combinación de ambas, en algunos indicadores de la relación suelo-planta en una plantación de morera establecida.
MATERIALES Y MÉTODOS
Ubicación geográfica. El estudio se realizó en áreas de la Estación Experimental de Pastos y Forrajes Indio Hatuey, del municipio Perico provincia de Matanzas, Cuba, situada entre los 22° 48' 7'' de latitud Norte y los 81° 2' de longitud Oeste, a 19,01 msnm (Academia de Ciencias de Cuba, 1989).
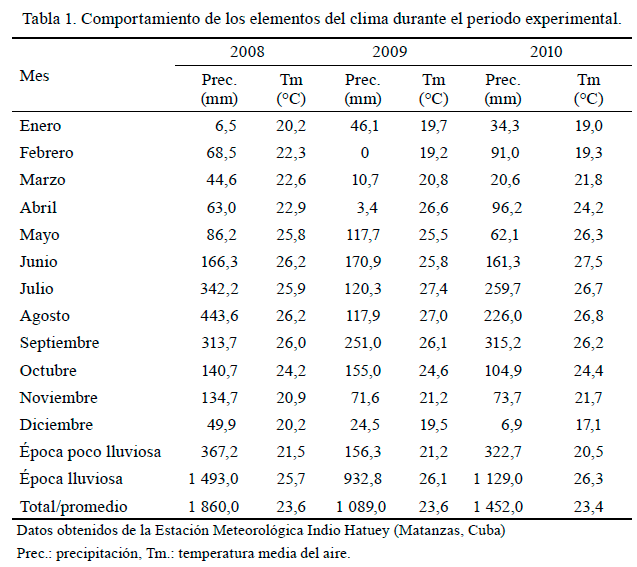
Características edafoclimáticas. Durante el periodo experimental (noviembre de 2008-noviembre de 2010) la precipitación se caracterizó por su variabilidad, con acumulados anuales que variaron entre 1 089 y 1 860 mm (tabla 1). El 18,77 % de las precipitaciones correspondieron a la época poco lluviosa. Los valores promedio de temperatura oscilaron entre 21,07 °C en la época poco lluviosa y 26,04 °C en la lluviosa.
El suelo es de topografía llana y se clasifica como Ferralítico Rojo hidratado (Hernández et al., 2003), con buen drenaje superficial e interno. La composición química se muestra en la tabla 2.
Origen y características de los recursos genéticos empleados. Las especies vegetales utilizadas en la investigación fueron M. alba L. var. tigreada, como cultivo principal, y C. ensiformis, como medio de multiplicación de los HMA. El producto comercial empleado para inocular los HMA fue EcoMic®, cepa Glomus cubense (Fernández et al., 2001).
Procedimiento. La morera tenía cuatro años de edad al iniciar el experimento. El marco de plantación fue de 26 666 plantas ha-1, con un arreglo espacial en doble surco (1,0 x 0,50 x 0,50 m). La dimensión de las parcelas fue de 24 m2 y los surcos se encontraban orientados en sentido este-oeste. Se asumió el criterio de no aplicar riego, de acuerdo con la situación real de la mayoría de las explotaciones agropecuarias comerciales. Durante los primeros 15 días de mayo y de noviembre, según el tratamiento establecido, se aplicó fertilización química de forma manual, localmente por planta. A continuación se inoculó con HMA directamente al suelo o a través de semillas de canavalia, recubiertas y sembradas entre los surcos de morera, a una distancia entre plantas de 40 cm y a 50 cm de los surcos de morera.
A los 90 días de iniciada la época poco lluviosa y a los 60 días de la lluviosa, se cortaron las plantas de canavalia y se arroparon sobre el surco de morera.
Fertilización química. Se emplearon tres dosis de fertilizante químico (F): 1) testigo sin fertilización (F0); 2) 100-50-50 kg de N, P y K ha-1 por época (F1); y 3) 200-100-100 kg de N, P y K ha-1 por época (F2). Las fuentes de fertilizante fueron: urea, superfosfato simple y cloruro de potasio.
Además, se analizó el perfil del suelo en la variable densidad radical. Las profundidades evaluadas fueron: 0-20, 20-40, 40-60 y 60-80 cm.
Diseño experimental. El diseño fue de cuatro bloques al azar, con arreglo factorial 3 x 2.
Tratamientos establecidos en el análisis bifactorial
F0: sin HMA ni fertilización
CeHMAF0: HMA con canavalia intercalada y sin fertilización
F1: 100-50-50 kg de N, P y K ha-1 por época
CeHMAF1: HMA vía canavalia, complementada con 100-50-50 kg de N, P y K ha-1 por época
F2: 200-100-100 kg de N, P y K ha-1 por época
CeHMAF2: HMA vía canavalia, complementada con 200-100-100 kg de N, P y K ha-1 por época
Indicadores evaluados
Densidad radical (expresada en miligramos por cada 50 g de suelo) de la morera en el perfil del suelo, cada 20 cm y hasta 80 cm de profundidad, al final del periodo experimental. Esta se determinó a partir de la extracción de bloques de suelo de 25 x 25 x 20 cm, a ambos lados de las plantas seleccionadas, que conformaron 1/3 del volumen de suelo correspondiente a la superficie vital del cultivo hasta los 80 cm de profundidad. El sistema radical formado por raíces y raicillas se separó a través de un tamiz, por vía húmeda, y se pesó en base seca. El tamaño de muestra fue de dos plantas por parcela.
Conteo de esporas a 20 cm de profundidad (número de esporas por 50 g de suelo). Durante dos años se estudió la dinámica de multiplicación de estructuras infectivas (esporas) a los 15 días, después del primer y el segundo corte de la época poco lluviosa y del primer corte de la época lluviosa, en los tratamientos CeHMAF0 y F0. Además, se evaluó al final del periodo experimental, en la época lluviosa, a los 150 días de aplicados los tratamientos. El procedimiento para la evaluación consistió en la toma de muestras de 50 g de suelo, tamizado y decantado por vía húmeda de los propágulos del hongo, según el método de extracción descrito por Gerdeman y Nicholson (1963). Las esporas se colectaron en una malla de 40 µm. Después se separaron por centrifugación con sacarosa y Tween 80, y se observaron en un estereomicroscopio óptico (20-40 x). La muestra fue de dos plantas por parcela.
La colonización fúngica (%) y la densidad visual de la colonización (%) a 20 cm de profundidad se determinaron al final del periodo experimental, en la época lluviosa, a los 150 días de aplicados los tratamientos. Para ello, se motearon las raíces de las plantas colectadas hasta 20 cm de profundidad. A continuación se lavaron con agua corriente para eliminar todo el suelo y se secaron al aire. Después se tomaron las raicillas más finas y se desmenuzaron. Se pesaron, aproximadamente, 200 mg de raicillas, que fueron secadas a 70 °C, y se tiñeron de acuerdo con el método descrito por Phillips y Hayman (1970). La evaluación se realizó según el método de los interceptos, desarrollado por Giovanetti y Mosse (1980), para determinar el porcentaje de colonización micorrízica o la frecuencia de colonización, a partir de la fórmula:
El porcentaje de densidad visual (% DV) se determinó por el método de Trouvelot et al. (1986), mediante la cual se evaluó la ocupación fúngica de cada intercepto y se le asignó un nivel, que se describe a continuación:
Donde:
Z: número de interceptos contados en cada nivel.
A: resultado de la multiplicación de Z por el porcentaje de ocupación observada.
La evaluación de los indicadores de la simbiosis micorrízica se realizó al final de la época lluviosa, a los 150 días de la inoculación con HMA, porque en esta época se expresa mejor el comportamiento micotrópico de la morera. Además, se consideró 150 días como un periodo que permite evaluar la permanencia de los propágulos infectivos en el suelo y la eficiencia de la colonización en fecha previa a la reinoculación.
Extracción de nitrógeno a través de la biomasa comestible de morera. La extracción de nitrógeno (N) se calculó durante dos años (kg ha-1 por época), a partir de los datos de la masa seca y su correspondiente concentración, mediante la fórmula:
Extracción de N = [Masa seca x % de N en hojas y tallos tiernos] /100
Procesamiento estadístico. El análisis de los resultados se realizó por época, teniendo en consideración los resultados de Martín (2004), quien demostró la estacionalidad de la morera. Se comprobaron los supuestos de normalidad de los errores, así como la homogeneidad de la varianza. Asimismo, se transformaron las variables en «x para los datos porcentuales (colonización fúngica y densidad visual) o log x para el número de esporas de HMA. Se empleó un modelo lineal general que incluyó los efectos estudiados y sus interacciones (Yijk = ì + Fi+ inoculación de HMAj + bloquek + eijkl.). Para la comparación de las medias se utilizó la dócima de Duncan (1955), para p ≤ 0,05. El paquete estadístico empleado fue InfoStat versión libre (InfoStat, 2004).
RESULTADOS Y DISCUSIÓN
En las figuras 1 y 2 se muestra el efecto significativo (p≤0,05) de las alternativas de inoculación y el perfil del suelo, como factores independientes, en la densidad radical. Las alternativas con HMA inoculados a través de canavalia intercalada fueron notablemente superiores a F0 y F1. Hubo tendencia al aumento en el volumen de raíces en la medida que se complementó la inoculación de HMA con fertilizante, aunque sin diferencias significativas. Ello se fundamenta en el hecho de que las hifas del hongo capturan el nitrógeno simple de la materia orgánica en descomposición, que se encuentra en el suelo, y facilitan el acceso a los macro- y micronutrientes disponibles, lo que estimula el crecimiento de la planta, y, en presencia de fertilizante, propicia un desarrollo intenso de las raíces (Hodge, 2002).
En el tratamiento CeHMAF0, la ausencia de fertilización química debió ser compensada por la absorción de los elementos minerales a través de las estructuras HMA, lo que exigió un mayor desarrollo radical. Esto fue estimulado, además, por la presencia de la canavalia (sembrada a baja densidad). Entry et al. (2002) afirmaron que las raíces micorrizadas son capaces de obtener más nutrientes que las que no lo están, debido a que las hifas exploran un mayor volumen de suelo. Ello tiene gran importancia en las condiciones de este estudio, ya que el contenido de materia orgánica en el suelo fue medio, y el potasio intercambiable, bajo. Paneque y Calaña (2001) consideraron que los valores inferiores a 0,15 o entre 0,15 y 0,30 cmol kg-1 de potasio indican baja presencia de este catión en el suelo. En cuanto al contenido de materia orgánica, los autores calificaron como medio un 3 % de MO.
Con respecto a la incidencia del perfil del suelo en la densidad radical (fig. 2), la tendencia fue a disminuir en las capas más profundas. El volumen promedio de raíces fue significativamente mayor en 0-20 cm, y no existieron diferencias notables en 20-40 y 40-60 cm, ni en 40-60 y 60-80 cm.
Es conocido que la densidad de las raíces se presenta en gradientes típicos entre la capa superficial del suelo y el subsuelo. La alta densidad de las raíces en las capas superficiales es causada, fundamentalmente, por condiciones físicas, químicas y biológicas más favorables, y el logaritmo de la densidad de las raíces disminuye linealmente con el incremento de la profundidad (Rivera, 1987).
Se debe reconocer que, a pesar de la menor densidad de raíces en el subsuelo, la absorción de nutrientes procedentes de esta zona puede ser considerable. La absorción desde el subsuelo también es significativa para los nutrientes minerales, tales como: el nitrato de amonio, el magnesio, el potasio y el fósforo. La importancia relativa del suministro a partir del subsuelo depende, entre otros aspectos, de la densidad de las raíces en esta zona (Hernández, 2001).
En
la figura 3 s
e
describe la dinámica de multiplicación de estructuras de HMA entre
0-20 cm. El contenido de propágulos nativos en la rizosfera varió
entre 75 y 434 esporas en 50 g de suelo, en el tratamiento F0, lo que se considera
entre medio y bajo de acuerdo con los criterios de Rivera y Fernández (2003).
Estos autores clasifican la densidad como baja cuando es menor de 200 esporas
en 50 g de suelo; media, entre 300-500; y alta, cuando supera las 600 esporas
en 50 g de suelo. Dichos criterios permiten considerar efectivo el tratamiento
CeHMAF0, ya que en casi todos los cortes el número de estructuras infectivas
fue superior a 400 esporas en 50 g de suelo, con relevancia en el primer corte
de la época lluviosa del primer año, el cual alcanzó valores
de 917. En este sentido, la diferencia con respecto a F0 fue notable y, además,
constituye un indicador de que la morera es una especie micótrofa.
La disminución
de la población de esporas en el mes de febrero del primer año
fue considerable, tanto en F0 como en CeHMAF0, lo que demostró las limitaciones
en la capacidad de multiplicación de estructuras infectivas durante los
meses más secos.
Al
respecto se conoce que los HMA son simbiontes obligatorios y que su distribución
en los suelos cultivados está fuertemente influenciada por la vegetación,
las variables del clima y las características del suelo. La población
de esporas de HMA se incrementa significativamente con el aumento del número
de especies vegetales presentes en un sistema agrícola (Chen
et al., 2004), en contraste con las plantaciones de morera en monocultivo.
Peña et al. (2006), al realizar el análisis de frecuencia de aparición
de esporas nativas de HMA por tipo de cobertura, encontraron que los sistemas
agroforestales, seguidos por el bosque natural, presentaron los mayores promedios
de colonización radical (por encima del 30 %). Por el contrario, las
coberturas de parcelas, así como el monocultivo y los potreros presentaron
los valores de colonización radical más bajos. En otros estudios,
estos autores demostraron que la simbiosis micorrízica arbuscular fue
más efectiva en ecosistemas con coberturas altamente heterogéneas;
de ahí la importancia de promover sistemas de producción agrícola
biodiversos (Peña
et
al., 2007), que minimicen la fragilidad de las plantaciones de monocultivo,
como es el caso de la morera.
En la tabla 3 se
presentan los resultados de la multiplicación de propágulos infectivos
en el momento de la inoculación y después de 150 días de
aplicados los tratamientos. El contenido de propágulos nativos en la
rizosfera (F0) fue bajo.
Hubo un efecto significativo de todos los tratamientos en la multiplicación de esporas después de 150 días de aplicados, en la época de lluvia. La densidad visual de las raíces de morera fue notablemente mayor en el tratamiento CeHMAF1.
La mayor densidad visual y la colonización en F1, en comparación con la dosis máxima (F2), confirma lo señalado por Rivera y Fernández (2003) acerca del efecto negativo de las altas dosis de fertilizante químico sobre la simbiosis micorrízica. Asimismo, la disminución de su efectividad es una consecuencia del mal funcionamiento de la estructuras o de la inhibición de la simbiosis.
El
alto suministro de fósforo puede provocar un desbalance entre este elemento,
el nitrógeno y el potasio, que estimula el crecimiento radical pero inhibe
el funcionamiento fúngico. Por el contrario, si la disponibilidad de
fósforo es baja o nula, no funciona adecuadamente la simbiosis, y la
efectividad de la inoculación es baja (Mohammad
et al., 2004). Se estima que la asociación entre el hospedante y los
HMA consume entre 5 y 10 % de los productos de la fotosíntesis. Este
costo se compensa si la planta se encuentra en condiciones subóptimas
de suministro de nutrientes (Siqueira et al.
,
2010).
Los valores de
colonización por debajo de 25 %, a los 150 días del inicio de
la época de lluvia, indicaron la necesidad de introducir artificialmente
HMA, a través de la inoculación al inicio y al final; y aumentar
así los efectos de la simbiosis, tanto en la época lluviosa como
en la poco lluviosa.
En este sentido, en varios trabajos se ha comprobado la influencia del tipo de suelo y la estacionalidad sobre el funcionamiento micorrízico. En suelos con buen drenaje, los mayores porcentajes de colonización se encontraron en la estación lluviosa (Apple et al., 2005; De Oliveira y de Oliveira, 2005; Becerra et al., 2007); mientras que en la medida que los suelos fueron menos fértiles se hizo necesaria una mayor cantidad de estructuras fúngicas, para garantizar el funcionamiento adecuado de la simbiosis (Azcón et al., 2003).
En
relación con las extracciones de nitrógeno a través de
la biomasa comestible de morera (tabla 4), se observó que las mayores
extracciones coincidieron con los tratamientos de mayor aporte de nutrientes.
Los valores más altos oscilaron entre 245,68
y 336,75 kg ha-1 de N en la época de lluvia y entre 80,63
y 108,08 kg ha-1de N
en
la poco lluviosa. Los tratamientos CeHMAF1, F2 y CeHMAF2 no difirieron entre sí
en el primer año de evaluación y fueron significativamente superiores
al resto. En el segundo año las mayores extracciones se obtuvieron en CeHMAF1.
Los resultados
evidenciaron la conveniencia de la inoculación de HMA a través
de canavalia intercalada, complementada con dosis de fertilizante por debajo
de lo óptimo, ya que el volumen de nitrógeno extraído en
esta condición no difirió de la dosis máxima. Además,
la fertilización fue más eficiente.
De manera general, se evidenció la alta capacidad extractiva de la morera en todos los tratamientos, lo que se corresponde con la literatura internacional. En este sentido, algunos datos compilados por Ito y Takagishi (1990) demostraron que en plantaciones de morera con rendimiento de 24,8 t ha-1 de biomasa foliar por año, el total de nitrógeno extraído desde el suelo ascendió a 242 kg ha-1 por año; de estos, 206 kg ha-1 por año fueron exportados de la plantación a través de las hojas.
CONCLUSIONES
Resulta factible el uso de la canavalia como medio de multiplicación de propágulos infectivos de HMA, dentro de las plantaciones de morera.
La mayor distribución de raíces de M. alba L. en el suelo estuvo asociada de manera positiva a la inoculación de HMA a través de canavalia, complementada con fertilización química.
La morera posee carácter micótrofo, y es efectiva la simbiosis micorrízica después de 150 días de aplicados los tratamientos en la época lluviosa.
Es necesario fertilizar con NPK, a razón de 100-50-50 kg ha-1 por época, al inicio y al final de la estación lluviosa.
En la condición anterior se garantizó una estrecha relación suelo-planta a través del crecimiento radical, que favoreció la simbiosis micorrízica como indicador del estado de funcionamiento micotrópico del cultivo y el suelo, y las altas extracciones de nitrógeno como respuesta del cultivo a la capacidad productiva del suelo.
REFERENCIAS BIBLIOGRÁFICAS
1. Academia de Ciencias de Cuba. Nuevo Atlas Nacional de Cuba. Instituto Cubano de Geodesia y Cartografía. La Habana, Cuba. p. 41. 1989.
2. Apple, M.E.; Thee, C.I.; Smith, V.L.; Cogar, C.R.; Wells, C.E. & Nowak, R.S. Arbuscular mycorrhizal colonization of Larrea tridentate and Ambrosia dumosa roots varies with precipitation and season in the Mojave Desert. Symbiosis. 39: 1 5. 2005.
3. Azcón, R.; Ambrosano, E. & Charest, C. Nutrient acquisition in mycorrhizal lettuce plants under different phosphorus and nitrogen concentration. Plant Science. 165: 1137 1145. 2003.
4. Becerra, A.G.; Arrigo, N.M.; Bartoloni, N.; Domínguez, L.S. & Cofré, M.N. Arbuscular mycorrhizal colonization of Alnus acuminata Kunth in northwestern Argentina in relation to season and soil parameters. Revista Ciencia del Suelo. Volumen 25 No. 1: 713. 2007.
5. Calderón, Maida & González, P. J. Respuesta del pasto guinea (Panicum maximum cv. Likoni) cultivado en suelo ferralítico rojo lixiviado a la inoculación de hongos micorrízicos arbusculares. Cultivos Tropicales, Vol. 28:33. 2007.
6. Chen, X.; Tang, J.; Fang, Z. & Shimizu, K. Effects of weeds communities with various species numbers on soil features in a subtropical orchard ecosystem. Agriculture, Ecosystem and Environment. 102 (3): 377388. 2004.
7. de Oliveira, A.N. & de Oliveira, L.A. Seasonal dynamics of arbuscular mycorrhizal fungi in plants of Theobroma grandiflorum Schum and Paullina cupana Mart. of an agroforestry system in Central Amazonia, Amazonas State, Brazil. Brazilian Journal of Microbiology. 36: 262270. 2005.
8. Duncan, D. B. Multiple ranges and multiple F. test. Biometrics. 11:1 1955.
9. Entry, I.A.; Rygiewicz, P.T.; Watrud, L.S. & Donnelly, P.K. Influence of adverse soil conditions on the formation and function of arbuscular mycorrhizas. Advances in Environmental Research. 7: 123138. 2002.
10. Fernández, F., Gómez, R., Martínez, M.A. & de la Noval, B.M. Producto inoculante micorrizógeno. Patente No. 22 641. Cuba. 2001
11. Gerdemann, J.W. & Nicholson, T.H. Spore of mycorrhizae endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc., 46: 235-244. 1963.
12. Giovannetti, M. & Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 84: 489500. 1980.
13. González, P.J.; Plana, R.; Rivera, R.; Fernández, F. & Arbola, J. Efectos de la inoculación de hongos micorrízicos arbusculares en pastos del género Brachiaria, cultivados en suelo pardo mullido. Revista Cubana de Ciencia Agrícola, Vol. 42:101. 2008.
14. Hernández, A.; Ascanio, M.; Cabrera, A.; Morales, M.; Medina, N. & Rivero, L. Nuevos aportes a la clasificación genética de suelos en el ámbito nacional e internacional. Instituto de Suelos. Ministerio de la Agricultura. AGRINFOR. La Habana, Cuba. 145p. 2003.
15. Hernández, R. 2001. Nutrición de plantas. Libro de Botánica on line. Copyright © 2001 - Version 2.0. 53Pp. http://www.forest.ula.ve/rubenhg. [5 de junio 2013].
16. Hodge, A. Microbial Ecology of the Arbuscular Mycorrhiza. FEMS Microbiol Ecol.32:91-96. 2002.
17. InfoStat. Versión 2004. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. 2004.
18. Ito. M. & Takagishi, H. Mulberry. The Fertilization Research Foundation, Tokyo, Japan, Japan International Cooperation System, Tokyo, Japan. (On line).<http://216.239.59.104/search?q=cache:N9r99CXi8moJ:www.fertilizer.org/ifa /publicat/html/pubman/mulberry.pdf+morus+alba+density&hl=es> 1997. [11/03/2005]
19. Martín, G. J. Evaluación de los factores agronómicos y sus efectos en el rendimiento y la composición bromatológica de Morus alba Linn. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. Universidad de Matanzas Camilo Cienfuegos. Matanzas, Cuba. 95 p. 2004.
20. Martín, G., Hernández, I. Milera, M. García, F. & González, T. Agronomic studies with mulberry in Cuba. Mulberry for animal production. FAO Bulletin 147.p 103-113. ISSN 0254- 6019. 2002.
21. Martín, Gloria M. Manejo de la inoculación micorrízica arbuscular, la Canavalia ensiformis y la fertilización nitrogenada en plantas de maíz (Zea mays) cultivadas sobre suelos ferralíticos fojos de La Habana. Tesis en opción al Grado Científico de Doctor en Ciencias Agrícolas. Instituto Nacional de Ciencias Agrícolas. 131 p. 2009.
22. Mohammad, A.; Mitra, B. & Khan, A.G. Effects of shearedroot inoculum of Glomus intraradices on wheat grown at different phosphorus levels in the field. Agriculture, ecosystems and environment. 103 (1): 245 249. 2004.
23. Noda Yolai & Castañeda Lisset. Efecto del EcoMic® en la emergencia de plántulas de Jatropha curcas. Pastos y Forrajes 35 (4): 401-406. 2012.
24. Oropesa, Katerine, Pentón, Gertrudis & Martín, G. J. Efecto de la fertilización biológica y/o mineral en la producción de forraje de morera (Morus alba L.) (Nota técnica). Pastos y Forrajes 34(3):295-302. 2011.
25. Paneque, V.M. & Calaña, J.M. La fertilización de los cultivos: aspectos teóricos prácticos para su recomendación. Folleto impreso. Curso de postgrado. Instituto Nacional de Ciencias Agrícolas. Departamento de biofertilizantes y nutrición de las plantas. San José de las Lajas, La Habana. pp 28. 2001.
26. Peña, C.P.; Cardona G.I.; Mazorra A.; Arguellez J. H. & Arcos A.L. Micorrizas arbusculares de la amazonia colombiana. Catalogo Ilustrado. Instituto Amazónico de Investigaciones Científicas SINCHI. 90 p. 2006.
27. Peña, C.P.; Cardona, G.I.; Arguelles, J.H. & Arcos, A.L. Micorrizas Arbusculares del Sur de la Amazonia colombiana y su relación con algunos factores fisicoquímicos y biológicos del suelo. Acta Amazónica. 37 (3): 327 326. 2007.
28. Phillips, D.M & Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 55. 158-161. 1970.
29. Rivera, R. Nutrición, fertilización y balance del fertilizante nitrogenado para el cafeto en un suelo ferralítico rojo compactado. Tesis presentada en opción al grado científico de candidato en ciencias agrícolas. La Habana, Cuba. 1987.
30. Rivera, R. & Fernández, K. Bases científicotécnicas para el manejo de los sistemas agrícolas micorrizados eficientemente. En: Rivera, R. & Fernández, K. Eds. Manejo efectivo de la simbiosis micorrízica, una vía hacia la agricultura sostenible. Estudio de caso: el Caribe. Instituto Nacional de Ciencias Agrícolas. La Habana., 166 p. 2003.
31. Siqueira, José Oswaldo; de Souza, Francisco A.; Cardoso, Eike J. B. N. & Tsai, Siu Mui. Micorrizas: 30 anos de pesquisas no Brasil. Ed. Lavras: UFLA. 716p. ISBN 978-85-87692-90-0. 2010.
32. Trouvelot, A.; Kough, J. & Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d'un systeme radiculaire. Recherche de methodes d'estimation ayantune signification functionnelle. Proc. st Eur. Symp. on Mycorrhizae: Physiological and genetical aspects of mycorrhizae, Dijón. INRA, Paris. 1986.
33. Villalba P. Estudio de la dosis adecuada de inoculación de Ecomic® a través del intercalamiento de abono verde de Canavalia ensiforme. Tesis de diploma en opción al título de Ingeniero agrónomo. Universidad de Matanzas Camilo Cienfuegos. 50 p. 2009.
Recibido el 10
de junio de 2013
Aceptado
el 20 de junio de 2014



{kind=link}
{kind=link}
{kind=link}
{kind=link}